Digitally reconstructing a slice (just a slice) of the rat neocortex allowed these scientists to study many slices of neocortex (using supercomputers) under different conditions such as sleep and when the brain is under stress! Then they saw that changing just one parameter, such as Ca++ ion concentration, can change the state of the brain. Then they theorized what if, in illness, the brain is stuck is the “wrong” mode! For example, when the fight or flight response is initiated through adrenocorticotrophic hormone, how do tunnel vision and aggression occur as a result of that? This is AMAZING! And this could be a valuable, valuable tool in studying mental illness. Imagine if they took a slice of my brain and reconstructed it digitally, then even a comparison to someone’s brain who doesn’t have bipolar d/o (BPD) would be enormously valuable. Also studying how my neurons fire, what are the ionic levels, how do these differ than “normal” neurons? How can we treat my neurons to better resemble normal neurons? This is an AMAZING technology that has been developed! Please, someone, start studying BPD with this and who knows, a cure may be imminent! I know people say “the sky’s the limit” all the time, but in this case, the sky really is the limit, in fact beyond the sky is the limit! Genius work, hope it affords us some tangible help and treatment options, and soon!
Summary: Scientists digitally reconstructed a slice of juvenile rat brain. This digital slice has over 31,000 neurons, 55 layers of cells and 207 different neuron subtypes. They have discovered nearly 40 million synapses and over 2,000 connections between each brain cell type. Wow!
The scientists then used super computers to see how these virtual brain slices work during sleep or when under stress! The principal scientist, Henry Markram said “It paves the way for predicting the location, numbers, and even the amount of ion currents flowing through all 40 million synapses.”
Once the reconstruction was complete, the investigators used powerful supercomputers to simulate the behavior of neurons under different conditions. Remarkably, the researchers found that, by slightly adjusting just one parameter, the level of calcium ions, they could produce broader patterns of circuit-level activity that could not be predicted based on features of the individual neurons. For instance, slow synchronous waves of neuronal activity, which have been observed in the brain during sleep, were triggered in their simulations, suggesting that neural circuits may be able to switch into different “states” that could underlie important behaviors.
“An analogy would be a computer processer that can reconfigure to focus on certain tasks,” Markram says. “The experiments suggest the existence of a spectrum of states, so this raises new types of questions, such as ‘what if you’re stuck in the wrong state?'” For instance, Markram suggests that the findings may open up new avenues for explaining how initiating the fight-or-flight response through the adrenocorticotropic hormone yields tunnel vision and aggression.
Article reference and part of article below: http://www.cell.com/cell/abstract/S0092-8674(15)01191-5?_returnURL=http%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS0092867415011915%3Fshowall%3Dtrue
Reconstruction and Simulation of Neocortical Microcircuitry Introduction
Since Santiago Ramón y Cajal’s seminal work on the neocortex (DeFelipe and Jones, 1988, Ramón y Cajal, 1909, 1911), a vast number of studies have attempted to unravel its multiple levels of anatomical organization (types of neurons, synaptic connections, layering, afferent and efferent projections within and between neocortical regions, etc.) and functional properties (neuronal response characteristics, synaptic responses and plasticity, receptive fields, functional neocortical columns, emergent activity maps, interactions between neocortical regions, etc.). However, there are still large gaps in our knowledge, especially concerning the anatomical and physiological organization of the neocortex at the cellular and synaptic levels.
Specifically, while neurons have been classified in terms of their electrophysiological behaviors (Connors and Gutnick, 1990, Kasper et al., 1994,McCormick et al., 1985), expression of different calcium-binding proteins and neuropeptides (Celio, 1986, DeFelipe, 1993, Gonchar and Burkhalter, 1997,Kawaguchi and Kubota, 1997, Toledo-Rodriguez et al., 2005) and morphological features (Kisvárday et al., 1985, Larkman, 1991a, Tamás et al., 1998, Wang et al., 2002), there is still no consensus on an objective and comprehensive classification of neuron types. Although the distribution of protein and genetic markers for different neurons (Grange et al., 2014, Hendry et al., 1989,Kawaguchi and Kubota, 1997, Meyer et al., 2002, Toledo-Rodriguez et al., 2004) and the relative proportions of some morphologically and electrically classified neurons (Beaulieu and Colonnier, 1983, Cauli et al., 1997, Hendry et al., 1984,Meyer et al., 2010a, Rudy et al., 2011) have been described, we lack a comprehensive view of the number of each type of neuron in each layer. Since the advent of paired recording techniques, several studies have characterized the anatomical and physiological properties of synaptic connections between some types of neurons (Cobb et al., 1997, Feldmeyer et al., 1999, Frick et al., 2008,Gupta et al., 2000, Mason et al., 1991, Reyes et al., 1998, Thomson et al., 1993), but a large proportion have yet to be studied. Although labeling with retrograde and anterograde tracers and trans-synaptic viral vectors, imaging with array tomography, and saturated reconstruction with electron microscopy have made it possible to begin mapping pre- and postsynaptic neurons for individual neocortical neurons (Boyd and Matsubara, 1991, Callaway, 2008, Glenn et al., 1982, Kasthuri et al., 2015, Killackey et al., 1983, Micheva and Smith, 2007,Micheva et al., 2010, Wickersham et al., 2007), we know neither the numbers and types of the pre- and postsynaptic neurons associated with any specific neuron type nor the numbers and locations of the synapses that they form with their immediate neighbors.
At a functional level, there have been many investigations of emergent behavior in neocortical slices (Cunningham et al., 2004, Mao et al., 2001, McCormick et al., 2003, Sanchez-Vives and McCormick, 2000, Yuste et al., 1997), correlated activity (Hasenstaub et al., 2005, Livingstone, 1996, Salinas and Sejnowski, 2001, Shu et al., 2003, Silberberg et al., 2004, Singer, 1993), and the functional impact of individual neurons across cortical layers (Sakata and Harris, 2009,Schroeder and Foxe, 2002, Silva et al., 1991, Steriade et al., 1993), as well as in vivo activity in somatosensory and other cortical areas (Chen et al., 2015,Klausberger et al., 2003, Leinekugel et al., 2002, Luczak et al., 2007, Reyes-Puerta et al., 2015, Wilson et al., 2012), However, we still lack an understanding of the cellular and synaptic mechanisms and the role of the different layers in the simplest of behaviors, such as correlated and uncorrelated single-neuron activity and, more generally, synchronous and asynchronous population activity. For example, it is known that different types of neurons are connected through synapses with different dynamics and strengths, strategically positioned at different locations on the neurons’ dendrites, somata, and axons, but the functional significance of this organization remains unclear. Computational approaches that abstract away this level of biological detail have not been able to explain the functional significance of such intricate cellular and synaptic organization. Although future experimental research will undoubtedly advance our knowledge, it is debatable whether experimental mapping alone can provide enough data to answer these questions.
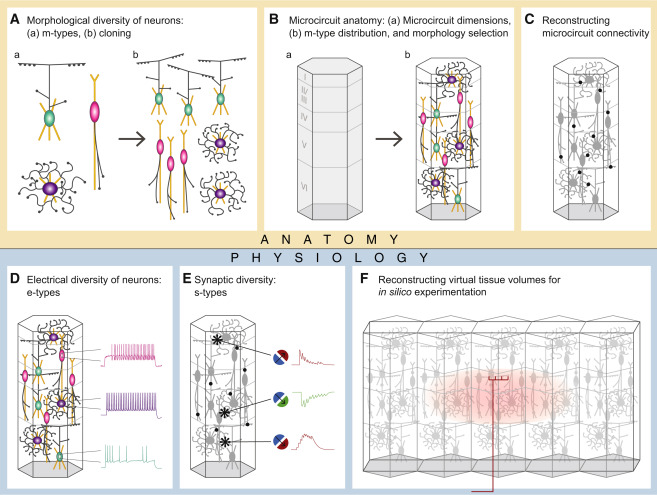
Here, we present a complementary algorithmic approach that reconstructs neuronal microcircuitry across all layers using available sparse data and that leverages biological principles and interdependencies between datasets to predict missing biological data. As a test case, we digitally reconstructed a small volume of tissue from layers 1 to 6 of the hind-limb somatosensory cortex of 2-week-old Wistar (Han) rat. This model system was chosen not only because it is one of the most comprehensively characterized in the neocortex, but also because experimental data on its cellular and synaptic organization are readily available and validation experiments are relatively easy to perform. In brief, we recorded and digitally reconstructed neurons from in vitro brain slices and classified the neurons in terms of well-established morphological types (m-types;Figure 1A), positioned the neurons in a digital volume of objectively defined dimensions according to experimentally based estimates of their layer specific densities (Figure 1B), and reconstructed the connectivity between the neurons (Figure 1C). Neurons were then classified into electrical types (e-types), using an extended version of the classification proposed in the Petilla convention (Ascoli et al., 2008), and models were produced that captured the characteristic electrical behavior of each type. (Figure 1D); similarly, synapses were modeled to capture the characteristic synaptic dynamics and kinetics of particular synapse types (s-types; Figure 1E). Finally, we constructed a virtual slice and reconstructed thalamic input using experimental data (Figure 1F; Meyer et al., 2010b).
This approach yielded a first-draft digital reconstruction of the microcircuitry, which was validated against a multitude of experimental datasets not used in the reconstruction. The results suggest that it is possible to obtain dense maps of neural microcircuitry without measuring every conceivable biological parameter and point to minimal datasets required, i.e., strategic data. Integrating complementary, albeit sparse, datasets also makes it possible to reconcile discrepancies in the literature, at least partially addressing the problem of data quality and reproducibility. Simulations exploring some of the emergent behaviors of the reconstructed microcircuitry reproduce a number of previous in vitro and in vivo findings and provide insights into the design and functioning of neocortical microcircuitry. The experimental data, the digital reconstruction, and the simulation results are available at the Neocortical Microcircuit Collaboration Portal (NMC Portal; https://bbp.epfl.ch/nmc-portal; see Ramaswamy et al., 2015).
